
SML1128
NMS-873
≥98% (HPLC)
別名:
3-[3-(シクロペンチルチオ-5-[[[2-メチル-4'-(メチルスルホニル)[1,1'-ビフェニル]-4-イル]オキシ]メチル]-4H-1,2,4-トリアゾール-4-イル]-ピリジン, 3-[3-シクロペンチルスルファニル-5-(4'-メタンスルホニル-2-メチルビフェニル-4-イルオキシメチル)-[1,2,4]トリアゾール-4-イル]-ピリジン
About This Item
おすすめの製品



アッセイ
≥98% (HPLC)
フォーム
powder
色
white to beige
溶解性
DMSO: 25 mg/mL, clear
保管温度
2-8°C
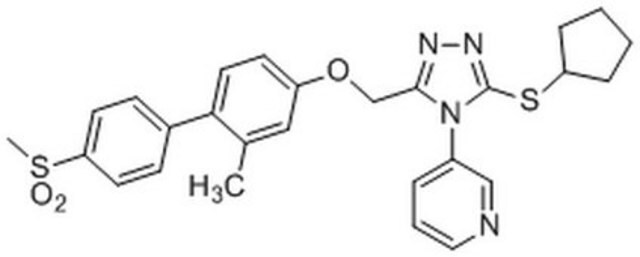
SMILES記法
CC1=CC(OCC2=NN=C(SC3CCCC3)N2C4=CN=CC=C4)=CC=C1C5=CC=C(S(C)(=O)=O)C=C5
InChI
1S/C27H28N4O3S2/c1-19-16-22(11-14-25(19)20-9-12-24(13-10-20)36(2,32)33)34-18-26-29-30-27(35-23-7-3-4-8-23)31(26)21-6-5-15-28-17-21/h5-6,9-17,23H,3-4,7-8,18H2,1-2H3
InChI Key
UJGTUKMAJVCBIS-UHFFFAOYSA-N
アプリケーション
生物化学的/生理学的作用
保管分類コード
11 - Combustible Solids
WGK
WGK 3
引火点(°F)
Not applicable
引火点(℃)
Not applicable
適用法令
試験研究用途を考慮した関連法令を主に挙げております。化学物質以外については、一部の情報のみ提供しています。 製品を安全かつ合法的に使用することは、使用者の義務です。最新情報により修正される場合があります。WEBの反映には時間を要することがあるため、適宜SDSをご参照ください。
Jan Code
SML1128-VAR:
SML1128-BULK:
SML1128-5MG:
SML1128-25MG:
最新バージョンのいずれかを選択してください:
この製品を見ている人はこちらもチェック


ライフサイエンス、有機合成、材料科学、クロマトグラフィー、分析など、あらゆる分野の研究に経験のあるメンバーがおります。.
製品に関するお問い合わせはこちら(テクニカルサービス)