Opis ogólny
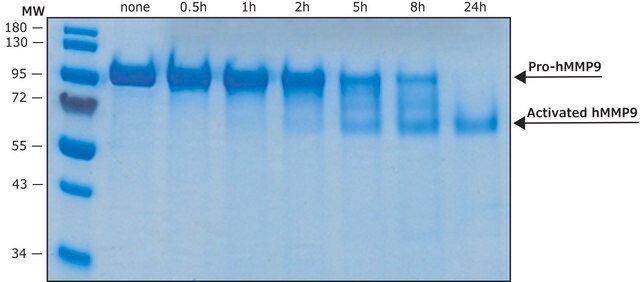
Rekombinowany ludzki pro-MMP-9 wyrażany w komórkach CHO. Obliczona masa cząsteczkowa wynosi ~77 kDa, ale pozorna masa cząsteczkowa wynosi ~92 kDa przez SDS-PAGE. Przydatny do immunoblottingu, testów rozszczepiania substratu i zymografii. Proenzym MMP-9 można mierzyć na podstawie jego zdolności do degradacji żelatyny w zymogramie. 0,5 ng enzymu wystarcza do uwidocznienia zdegradowanej żelatyny za pomocą niebieskiego barwnika Coomassie. Metaloproteinazy macierzy są członkami unikalnej rodziny enzymów proteolitycznych, które mają jon cynku w swoich miejscach aktywnych i mogą degradować kolageny, elastynę i inne składniki macierzy zewnątrzkomórkowej (ECM). Enzymy te są obecne u zdrowych osób i wykazano, że odgrywają ważną rolę w procesach takich jak gojenie się ran, ciąża i resorpcja kości. Jednak nadekspresja i aktywacja MMP została powiązana z szeregiem procesów patologicznych i stanów chorobowych związanych z rozpadem i przebudową ECM. Choroby te obejmują inwazję i przerzuty nowotworowe, reumatoidalne zapalenie stawów, choroby przyzębia i procesy naczyniowe, takie jak angiogeneza, hiperplazja błony wewnętrznej, miażdżyca i tętniaki. Niedawno MMP zostały powiązane z chorobami neurodegeneracyjnymi, takimi jak choroba Alzheimera i stwardnienie zanikowe boczne (ALS). Istnieją naturalne inhibitory MMP, tkankowe inhibitory metaloproteinaz macierzy (TIMP), a także opracowano syntetyczne inhibitory, które dają nadzieję na nowe możliwości leczenia tych chorób. Regulacja aktywności MMP może zachodzić na poziomie ekspresji genów, w tym transkrypcji i translacji, na poziomie aktywacji lub na poziomie hamowania przez TIMP. Zatem zaburzenia w każdym z tych punktów mogą teoretycznie prowadzić do zmian w obrocie ECM. Ekspresja jest ściśle kontrolowana przez cytokiny pro- i przeciwzapalne i/lub czynniki wzrostu, a po wyprodukowaniu enzymy są zwykle wydzielane jako nieaktywne zymogramy. Po aktywacji (usunięciu hamującego regionu propeptydowego cząsteczek) MMP podlegają kontroli przez lokalnie wytwarzane TIMP. Wszystkie MMP mogą być aktywowane in vitro związkami organomerkurialnymi (np. octanem 4-aminofenylomocznika), ale czynniki odpowiedzialne za fizjologiczną aktywację wszystkich MMP nie zostały jasno zdefiniowane. Liczne badania wskazują, że członkowie rodziny MMP mają zdolność do wzajemnej aktywacji. Aktywacja MMP in vivo jest prawdopodobnie krytycznym krokiem w zakresie ich biologicznego zachowania, ponieważ to właśnie ta aktywacja przechyli równowagę na korzyść degradacji ECM. Cechą charakterystyczną chorób z udziałem MMP wydaje się być brak równowagi stechiometrycznej między aktywnymi MMP i TIMP, co prowadzi do nadmiernego rozerwania tkanki i często jej degradacji. Określenie mechanizmów, które kontrolują tę nierównowagę, może otworzyć kilka ważnych opcji terapeutycznych specyficznych inhibitorów enzymów.
Zastosowanie
Immunoblotting (1 µg białka/pas)
Test rozszczepienia substratu (1 µg białka/pas, patrz odniesienia do aplikacji)
Zymografia (1 µg białka/tor, patrz referencje aplikacji)
Opakowanie
Należy zapoznać się z etykietą fiolki, aby uzyskać informacje o stężeniu specyficznym dla partii.
Ostrzeżenie
Toksyczność: Standardowa obsługa (A)
Definicja jednostki
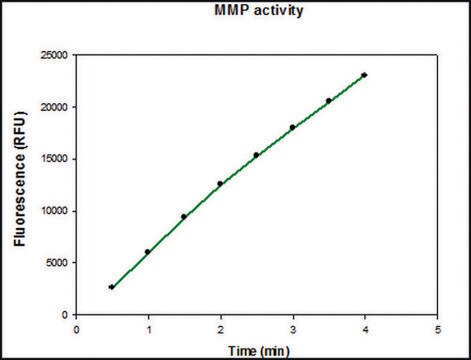
Aktywność specyficzną oznaczono przy użyciu 10 μM (7-metoksykumaryny-4-ylo)acetylo-Pro-Leu-Gly-Leu-(3-[2, 4-dinitrofenylo]-L-2, 3-diaminopropionylo)-Ala-Arg-NH₂ (wzbudzenie 320 nm, emisja 405 nm) i 20 ng enzymu w 100 μl 50 mM Tris-HCl, pH 7.5, 10 mM CaCl₂, 150 mM NaCl i 0,05% detergentu BRIJ®-35 w temperaturze pokojowej.
Postać fizyczna
W 150 mM NaCl, 50 mM Tris-HCl, 10 mM CaCl₂, 0,05% detergentu BRIJ®-35, pH 7,5.
Inne uwagi
Parsons, S.L., et al. 1997. Br. J. Surg.84, 160.
Backstrom, J.R., et al. 1996. J. Neuro.16, 7910.
Lim, G.P., et al. 1996. J Neurochem.67, 251.
Sang, Q.X., et al. 1995. Biochim. Biophys. Acta.1251, 99.
Kenagy, R.D. and Clowes, A.W. 1994. in Inhibition of Matrix Metalloproteinases: Therapeutic Potential. Greenwald, R.A. and Golub L.M., Eds.: 462-465.
Zempo, N., et al. 1994. J. Vasc. Surg.20, 209.
Birkedal-Hansen, H. 1993. J. Periodontol.64, 474.
Stetler-Stevenson, W.G., et al. 1993. FASEB J.7, 1434.
Delaisse, J-M. and Vaes, G. 1992. in Biology and Physiology of the Osteoclast. B.R. Rifkin & C.V. Gay, Eds.: 290-314.
Jeffrey, J.J. 1992. in Wound Healing: Biochemical and Clinical Aspects. R.F. Diegelmann and W.J. Lindblad, Eds.: 177-194.
Jeffrey, J.J. 1991. Semin. Perinatol.15, 118.
Liotta, L.A., et al. 1991. Cell64, 327.
Harris, E. 1990. N. Engl. J. Med.322, 1277.
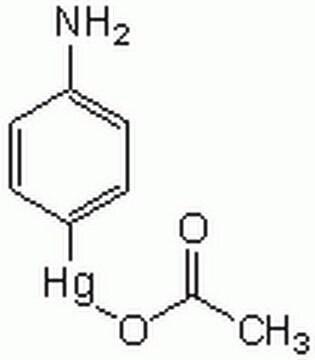
Proenzym MMP-9 może być mierzony poprzez jego zdolność do degradacji żelatyny w zymogramie. 0,5 ng enzymu wystarcza do uwidocznienia zdegradowanej żelatyny za pomocą niebieskiego barwnika coomassie. Aktywność specyficzna mierzona przy użyciu 10 µM (7-metoksykumaryny-4-ylo)acetylo-Pro-Leu-Gly-Leu-(3-[2, 4-dinitrofenylo]-L-2, 3-diaminopropionylo)-Ala-Arg-NH2 (wzbudzenie 320 nm, emisja 405 nm) i 20 ng enzymu w 100 µl 50 mM Tris-HCl, pH 7.5, 10 mM CaCl2, 150 mM NaCl i 0,05% Brij-35 w temperaturze pokojowej, wynosi > 1300 pmoli/min/µg. Aby aktywować proenzym MMP-9, należy przygotować koncentrat octanu p-aminofenylomocznika (APMA) w dimetylosulfotlenku (DMSO). Dodaj APMA do proenzymu MMP-9, aby uzyskać końcowe stężenie APMA 1 mM. Inkubować w temperaturze 37°C przez 16 do 24 godzin.
Informacje prawne
Brij is a registered trademark of Croda International PLC
CALBIOCHEM is a registered trademark of Merck KGaA, Darmstadt, Germany
Ta strona może zawierać tekst przetłumaczony maszynowo.