
17-10195
ProteoExtract® Cytoskeleton Enrichment and Isolation Kit
This kit provides cytoskeleton purification detergent buffers, sufficient for extraction from ten 100 mm culture dishes, that retain focal adhesion & actin-associated proteins while removing soluble cytoplasmic & nuclear proteins from the cell.
Iniciar sesiónpara Ver la Fijación de precios por contrato y de la organización
About This Item
Código UNSPSC:
12161503
eCl@ss:
32161000
Productos recomendados






fabricante / nombre comercial
Chemicon®
ProteoExtract®
técnicas
western blot: suitable
Condiciones de envío
dry ice
Descripción general
Read our application note in Nature Methods!
http://www.nature.com/app_notes/nmeth/2012/121007/pdf/an8624.pdf
(Click Here!)
Rediscovering the Actin Cytoskeleton: New techniques leading to discoveries in cell migration, Dr. Richard Klemke, Morris Cancer Center, UCSD, USA.
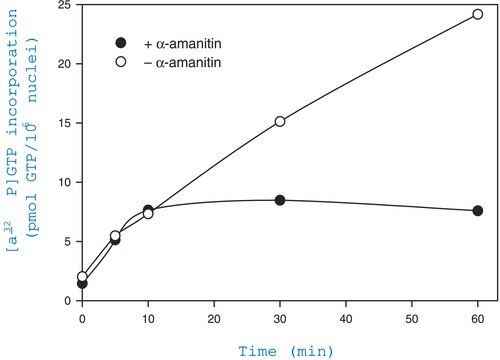
The actin cytoskeleton is a highly dynamic network composed of actin polymers and a large variety of associated proteins. The functions of the actin cytoskeleton is to mediate a variety of essential biological functions in all eukaryotic cells, including intra- and extra-cellular movement and structural
Support (Chen, C.S., et al., 2003; Frixione E., 2000). To perform these functions, the organization of the actin cytoskeleton must be tightly regulated both temporally and spatially. Many proteins associated with the actin cytoskeleton are thus likely targets of signaling pathways controlling actin assembly. Actin cytoskeleton assembly is regulated at multiple levels, including the organization of actin monomers (G-actin) into actin polymers and the superorganization of actin polymers into a filamentous network (F-actin – the major constituent of microfilaments) (Bretscher, A., et al., 1994). This superorganization of actin polymers into a filamentous network is mediated by actin side-binding or cross-linking proteins (Dubreuil, R. R., 1991; Matsudaira, P., 1991; Matsudaira, P., 1994). The actin cytoskeleton is a dynamic structure that rapidly changes shape and organization in response to stimuli and cell cycle progression. Therefore, a disruption of its normal regulation may lead to cell transformations and cancer. Transformed cells have been shown to contain less F-actin than untransformed cells and exhibit atypical coordination of F-actin levels throughout the cell cycle (Rao, J.Y., et al., 1990). Orientational distribution of actin filaments within a cell is, therefore, an important determinant of cellular shape and motility.
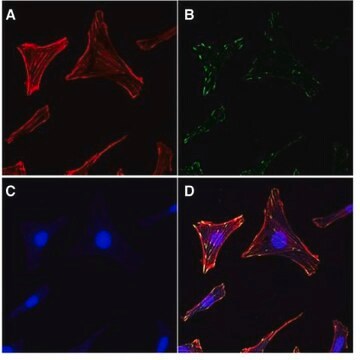
Focal adhesion and adherens junctions are membrane-associated complexes that serve as nucleation sites for actin filaments and as cross-linkers between the cell exterior, plasma membrane and actin cytoskeleton (Yamada, K.M., Geiger, B., 1997). The function of focal adhesions is structural, linking the ECM on the outside to the actin cytoskeleton on the inside. They are also sites of signal transduction, initiating signaling pathways in response to adhesion. Focal adhesions consist of integrin-type receptors that are attached to the extracellular matrix and are intracellularly associated with protein complexes containing vinculin (universal focal adhesion marker), talin, α-actinin, paxillin, tensin, zyxin and focal adhesion kinase (FAK) (Burridge, K., et al., 1990; Turner, C.E., Burridge, K., 1991).
Studying the proteins that associate with and regulate the actin cytoskeleton has been traditionally difficult because of the insolubility of the cytoskeleton in traditional detergents like Triton-X100. Work over the years has shown that many actin regulatory proteins/phospho-proteins upon activation move from the soluble cytoplasmic compartment to the insoluble actin cytoskeleton. The insolubility of these important proteins has made it difficult to study their biochemical changes, such as phosphorylation and nitrosylation, upon binding to actin. What is sorely needed is a cytoskeleton purification method that allows for the selective enrichment of cytoskeleton-associated proteins for detailed protein biochemical analyses. Such a method would provide the means to directly study this important pool of proteins in normal and diseased cytoskeletons.
http://www.nature.com/app_notes/nmeth/2012/121007/pdf/an8624.pdf
(Click Here!)
Rediscovering the Actin Cytoskeleton: New techniques leading to discoveries in cell migration, Dr. Richard Klemke, Morris Cancer Center, UCSD, USA.
The actin cytoskeleton is a highly dynamic network composed of actin polymers and a large variety of associated proteins. The functions of the actin cytoskeleton is to mediate a variety of essential biological functions in all eukaryotic cells, including intra- and extra-cellular movement and structural
Support (Chen, C.S., et al., 2003; Frixione E., 2000). To perform these functions, the organization of the actin cytoskeleton must be tightly regulated both temporally and spatially. Many proteins associated with the actin cytoskeleton are thus likely targets of signaling pathways controlling actin assembly. Actin cytoskeleton assembly is regulated at multiple levels, including the organization of actin monomers (G-actin) into actin polymers and the superorganization of actin polymers into a filamentous network (F-actin – the major constituent of microfilaments) (Bretscher, A., et al., 1994). This superorganization of actin polymers into a filamentous network is mediated by actin side-binding or cross-linking proteins (Dubreuil, R. R., 1991; Matsudaira, P., 1991; Matsudaira, P., 1994). The actin cytoskeleton is a dynamic structure that rapidly changes shape and organization in response to stimuli and cell cycle progression. Therefore, a disruption of its normal regulation may lead to cell transformations and cancer. Transformed cells have been shown to contain less F-actin than untransformed cells and exhibit atypical coordination of F-actin levels throughout the cell cycle (Rao, J.Y., et al., 1990). Orientational distribution of actin filaments within a cell is, therefore, an important determinant of cellular shape and motility.
Focal adhesion and adherens junctions are membrane-associated complexes that serve as nucleation sites for actin filaments and as cross-linkers between the cell exterior, plasma membrane and actin cytoskeleton (Yamada, K.M., Geiger, B., 1997). The function of focal adhesions is structural, linking the ECM on the outside to the actin cytoskeleton on the inside. They are also sites of signal transduction, initiating signaling pathways in response to adhesion. Focal adhesions consist of integrin-type receptors that are attached to the extracellular matrix and are intracellularly associated with protein complexes containing vinculin (universal focal adhesion marker), talin, α-actinin, paxillin, tensin, zyxin and focal adhesion kinase (FAK) (Burridge, K., et al., 1990; Turner, C.E., Burridge, K., 1991).
Studying the proteins that associate with and regulate the actin cytoskeleton has been traditionally difficult because of the insolubility of the cytoskeleton in traditional detergents like Triton-X100. Work over the years has shown that many actin regulatory proteins/phospho-proteins upon activation move from the soluble cytoplasmic compartment to the insoluble actin cytoskeleton. The insolubility of these important proteins has made it difficult to study their biochemical changes, such as phosphorylation and nitrosylation, upon binding to actin. What is sorely needed is a cytoskeleton purification method that allows for the selective enrichment of cytoskeleton-associated proteins for detailed protein biochemical analyses. Such a method would provide the means to directly study this important pool of proteins in normal and diseased cytoskeletons.
Aplicación
Research Category
Cell Structure
Cell Structure
Research Sub Category
Cytoskeleton
Cytoskeleton
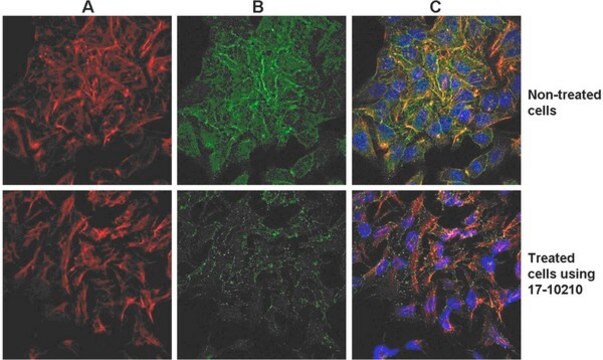
The EMD Millipore ProteoExtract® Cytoskeleton Enrichment and Isolation Kit provides cytoskeleton purification detergent buffers, sufficient for extraction from ten 100 mm culture dishes, that retain focal adhesion and actin-associated proteins while removing soluble cytoplasmic and nuclear proteins from the cell. Anti-Vimentin and anti-GAPDH antibodies are provided as markers for cytoskeleton and cytosol, respectively, in western blot analysis. This kit will greatly increase the ability to detect and study the low abundance actin-associated proteins which are typically masked in conventional methods of Western blotting analysis of whole cell lysates.
This kit provides cytoskeleton purification detergent buffers, sufficient for extraction from ten 100 mm culture dishes, that retain focal adhesion & actin-associated proteins while removing soluble cytoplasmic & nuclear proteins from the cell.
Envase
Enough Reagents for 15 Reactions/Isolations
Componentes
One bottle containing 1.9 mL of 10x Cellular Extraction Buffer
One bottle containing 3.5 mL of 20x Cytoskeleton Wash Buffer
One bottle containing 9 mL of ready to use Nuclear Extraction Buffer
Two vials each containing 100 μL of 1000x Protease Inhibitor Cocktail solution
One vial containing 150 μL of 1000x Sodium Orthovanadate solution
One bottle containing 3.8 mL of ready to use Cytoskeleton Solubilization Buffer
One vial containing 30 µL of 2000X goat anti-mouse HRP conjugate
One vial containing 50 µL of 1000x anti-GAPDH mouse monoclonal IgG1 antibody
One vial containing 100 µL of 500x anti-Vimentin mouse monoclonal IgG1 antibody
One bottle containing 3.5 mL of 20x Cytoskeleton Wash Buffer
One bottle containing 9 mL of ready to use Nuclear Extraction Buffer
Two vials each containing 100 μL of 1000x Protease Inhibitor Cocktail solution
One vial containing 150 μL of 1000x Sodium Orthovanadate solution
One bottle containing 3.8 mL of ready to use Cytoskeleton Solubilization Buffer
One vial containing 30 µL of 2000X goat anti-mouse HRP conjugate
One vial containing 50 µL of 1000x anti-GAPDH mouse monoclonal IgG1 antibody
One vial containing 100 µL of 500x anti-Vimentin mouse monoclonal IgG1 antibody
Almacenamiento y estabilidad
Maintain the unopened kit modules (17-10195-1 & 17-10195-2) at the indicated temperatures. The kit may be used for up to 4 months after receipt. Avoid repeated freeze/thaw cycles, and aliquot if necessary.
Información legal
CHEMICON is a registered trademark of Merck KGaA, Darmstadt, Germany
PROTEOEXTRACT is a registered trademark of Merck KGaA, Darmstadt, Germany
Cláusula de descargo de responsabilidad
Unless otherwise stated in our catalog or other company documentation accompanying the product(s), our products are intended for research use only and are not to be used for any other purpose, which includes but is not limited to, unauthorized commercial uses, in vitro diagnostic uses, ex vivo or in vivo therapeutic uses or any type of consumption or application to humans or animals.

Palabra de señalización
Warning
Frases de peligro
Consejos de prudencia
Clasificaciones de peligro
Acute Tox. 4 Oral - Aquatic Chronic 3 - Eye Irrit. 2 - Skin Irrit. 2 - Skin Sens. 1
Código de clase de almacenamiento
10 - Combustible liquids
Certificados de análisis (COA)
Busque Certificados de análisis (COA) introduciendo el número de lote del producto. Los números de lote se encuentran en la etiqueta del producto después de las palabras «Lot» o «Batch»
¿Ya tiene este producto?
Encuentre la documentación para los productos que ha comprado recientemente en la Biblioteca de documentos.
Viorica L Lastun et al.
The Journal of biological chemistry, 298(6), 101935-101935 (2022-04-19)
In metazoans, the architecture of the endoplasmic reticulum (ER) differs between cell types and undergoes major changes throughout the cell cycle and according to physiological needs. Although much is known about how the different ER morphologies are generated and maintained
Nuestro equipo de científicos tiene experiencia en todas las áreas de investigación: Ciencias de la vida, Ciencia de los materiales, Síntesis química, Cromatografía, Analítica y muchas otras.
Póngase en contacto con el Servicio técnico